熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
3. 結果與討論
3.1. 微觀體系實驗參數的變化
| 樣品 | 鈾 (Uranium) | K? | Na? | Ca2? | Mg2? | Mn2? | 總鐵 (Fe?????) | 生物可利用 Fe(III) |
|---|---|---|---|---|---|---|---|---|
| 原始沉積物 (g/kg) | 0.170 ± 0.005 | 13.38 ± 0.53 | 1.97 ± 0.09 | 55.38 ± 1.37 | 9.40 ± 0.32 | 0.72 ± 0.25 | 39.25 ± 1.31 | 0.35 ± 0.02 |
| 葡萄糖添加沉積物 (g/kg) | 0.174 ± 0.006 | 39.63 ± 1.82 | 6.71 ± 0.22 | 50.47 ± 1.42 | 3.92 ± 0.13 | 0.74 ± 0.22 | 45.10 ± 1.07 | 0.24 ± 0.01 |
| 乳酸添加沉積物 (g/kg) | 0.177 ± 0.006 | 39.38 ± 1.16 | 5.97 ± 0.31 | 53.40 ± 1.61 | 7.90 ± 0.35 | 0.71 ± 0.24 | 60.83 ± 1.76 | 0.29 ± 0.01 |
| 乙醇添加沉積物 (g/kg) | 0.176 ± 0.007 | 36.81 ± 0.99 | 2.21 ± 0.08 | 50.63 ± 1.55 | 6.17 ± 0.27 | 0.80 ± 0.34 | 72.75 ± 2.11 | 0.22 ± 0.01 |
| 原始地下水 (mg/L) | 1.020 ± 0.021 | 46.02 ± 1.42 | 170.80 ± 8.38 | 530.05 ± 22.36 | 41.75 ± 1.34 | UD | 0.28 ± 0.01 | - |
| 葡萄糖添加地下水 (mg/L) | 0.060 ± 0.018 | 46.12 ± 1.25 | 185.96 ± 6.22 | 266.95 ± 19.76 | 79.53 ± 1.65 | UD | 0.18 ± 0.01 | - |
| 乳酸添加地下水 (mg/L) | 0.030 ± 0.012 | 45.60 ± 1.17 | 200.40 ± 12.65 | 10.53 ± 0.42 | 79.06 ± 1.65 | UD | 0.22 ± 0.01 | - |
| 乙醇添加地下水 (mg/L) | 0.033 ± 0.009 | 44.98 ± 1.22 | 184.12 ± 7.35 | 230.63 ± 4.88 | 82.73 ± 2.97 | UD | 0.17 ± 0.01 | - |
表2顯示,除鈾和鈣外,地下水樣品中金屬離子的濃度變化不大。水相鈣濃度比其初始濃度下降了50%以上,水相鈾濃度比其初始濃度下降了90%以上。
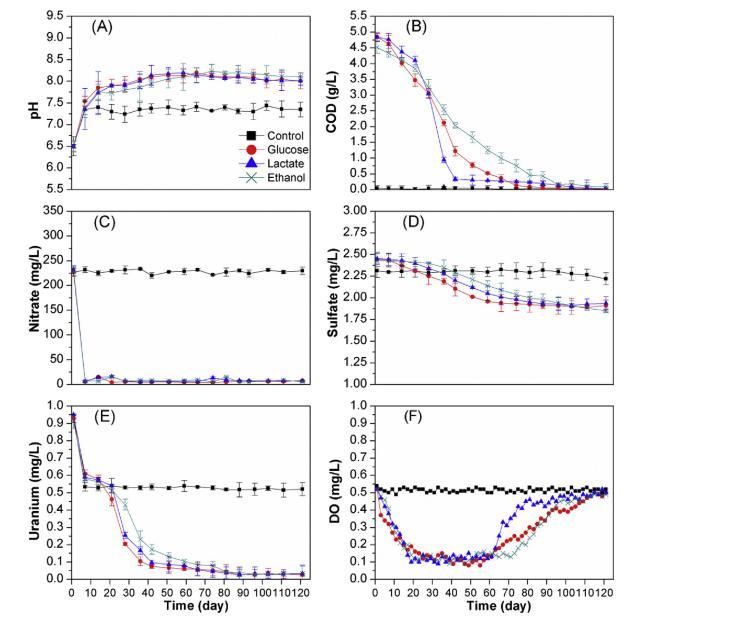
圖1. 培養期間微宇宙底部區域中(A) pH、(B) COD、(C) 硝酸鹽、(D) 硫酸鹽、(E) 鈾和(F) 氧溶解度的變化(初始濃度對應于第0天的濃度,所有數據點均為三次重復測量的平均值)。
圖1顯示,在培養的第一周,添加了電子供體的微觀體系中水相硝酸鹽濃度下降了90%以上,并在實驗剩余時間內保持耗竭狀態。當硝酸鹽從微觀體系中去除后,水相硫酸鹽濃度開始相繼下降,表明硫酸鹽還原菌受到了刺激。同時,添加了電子供體的微觀體系從低氧條件轉變為缺氧條件。添加了電子供體的微觀體系中的COD濃度被用作地下水中添加的電子供體濃度的指標。培養90天后,COD濃度變得檢測不到,表明地下水中的電子供體已耗盡。在培養的第一周,所有微觀體系中的pH均上升至7.2,這是由碳酸氫鈉與沉積物相互作用引起的。此后,對照中的pH變得穩定,而添加了電子供體的微觀體系中的pH則緩慢持續上升,直至電子供體耗盡。培養期間添加了電子供體的微觀體系中pH的升高源于硝酸鹽和硫酸鹽的還原。盡管Fe(III)礦物可能影響U(VI)的還原和U(IV)/U(VI)吸附,但在培養期間未監測Fe(III)的還原,因為沉積物中微生物可利用的Fe(III)含量和地下水中總鐵含量非常低。
3.2. 培養期間的U(VI)還原
在培養的第一周,所有微觀體系中的水相總溶解鈾均大幅下降,這是由于鈾被吸附到沉積物上。之后,添加了電子供體的微觀體系中水相總溶解鈾持續下降,而在對照中保持不變。在乙醇、乳酸和葡萄糖添加的微觀體系中,水相總溶解鈾分別在第96天、第88天和第96天降至0.030、0.025和0.026毫克/升,低于中國飲用水鈾最大污染物限值0.050毫克/升,甚至達到了美國環保署的飲用水最大污染物水平。盡管在第90天電子供體耗盡時,添加了電子供體的微觀體系重新變為低氧條件,但所有添加了電子供體的微觀體系中低濃度的水相鈾仍保持不變。
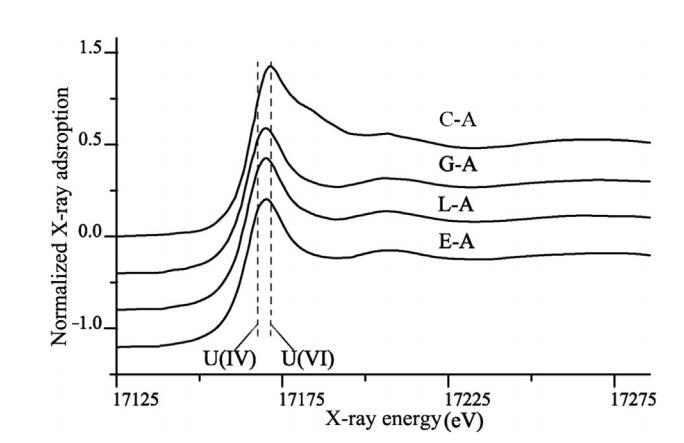
圖2.沉積物樣本在培養90天后的U L3-edge XANES 光譜(C-A代表第90天未經處理的微宇宙沉積物樣本)。
第90天沉積物樣品的U L3邊XANES測量結果(圖2)顯示,葡萄糖、乳酸和乙醇添加的微觀體系中分別約有42% ± 10%、45% ± 10%和43% ± 10%的鈾為U(IV),而對照中不存在U(IV)。添加了電子供體的微觀體系中U(IV)的存在證實了生物還原發生在微觀體系中,并且添加了電子供體的微觀體系中水相鈾的下降部分歸因于生物成因U(IV)的形成。在添加了不同電子供體的微觀體系之間,生物成因U(IV)的比例沒有觀察到明顯差異,這表明在低氧條件下,葡萄糖、乳酸和乙醇與在缺氧條件下一樣,都是有效的電子供體。
3.3. 溶解氧的變化
培養期間,對照中的DO濃度維持在0.5毫克/升,表明與對照在同一手套箱中培養的、添加了電子供體的微觀體系同樣處于低氧條件下。然而,當培養開始時,添加了電子供體的微觀體系中的DO濃度開始下降,培養數天后,缺氧條件取代了初始的低氧條件。當培養結束時電子供體耗盡,添加了電子供體的微觀體系中的DO濃度回升,微觀體系從缺氧狀態恢復到初始的低氧條件。培養期間,添加了電子供體的微觀體系中DO濃度的下降和缺氧區的形成源于受刺激微生物的代謝作用。同時,盡管恢復到低氧條件,添加了電子供體的微觀體系中的總溶解鈾并未顯著回升。
3.4. 微生物群落豐富度和多樣性
根據焦磷酸測序結果,計算了歸一化16S rRNA基因文庫的微生物群落豐富度和多樣性指數。根據數值大小,添加了電子供體的微觀體系的沉積物樣品中微生物群落豐富度的Chao和ACE指數排序如下:S-B > G-D > G-A > L-A > E-D > L-D > E-A。原始沉積物具有最高的微生物群落多樣性,但來自添加了電子供體的微觀體系的沉積物多樣性較低,這與已報道的結果一致。電子供體的添加可能對與U(VI)生物還原相關的微生物起到富集作用,導致與U(VI)生物還原無關的其他微生物比例相對下降。稀疏曲線顯示的微生物群落豐富度特征與Chao和ACE指數結果相同。
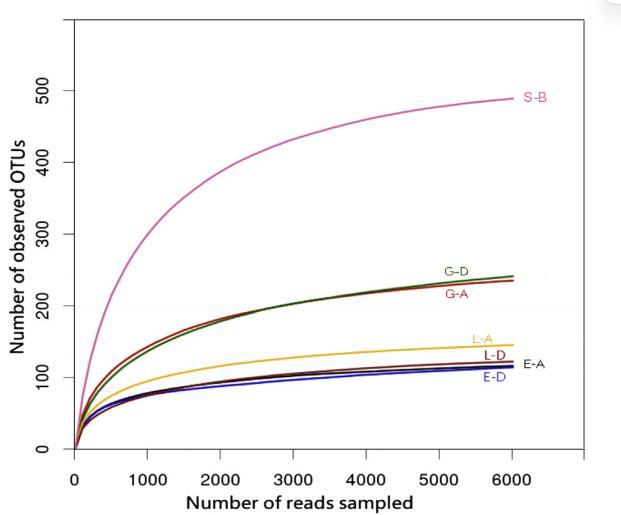
圖3. 沉積物樣本中觀察到的操作分類單元(OTUs)的稀釋曲線。
表3:沉積物樣品在門水平上的微生物群落豐富度(相對豐度 ≥ 1%)| 分類單元 (Taxon) | E-D (%) | E-A (%) | L-D (%) | L-A (%) | G-D (%) | G-A (%) | S-B (%) |
|---|---|---|---|---|---|---|---|
| 厚壁菌門 (Firmicutes) | 48 | 42 | 72 | 55 | 16 | 20 | 1 |
| 擬桿菌門 (Bacteroidetes) | 0 | 0 | 4 | 10 | 19 | 25 | 0 |
| 放線菌門 (Actinobacteria) | 0 | 0 | 0 | 0 | 0 | 0 | 31 |
| 酸桿菌門 (Acidobacteria) | 0 | 0 | 0 | 0 | 0 | 0 | 10 |
| 芽單胞菌門 (Gemmatimonadetes) | 0 | 0 | 0 | 0 | 0 | 0 | 8 |
| 變形菌門-α綱 (Alphaproteobacteria) | 1 | 4 | 5 | 14 | 8 | 3 | 22 |
| 變形菌門-β綱 (Betaproteobacteria) | 25 | 24 | 9 | 6 | 6 | 7 | 7 |
| 變形菌門-δ綱 (Deltaproteobacteria) | 1 | 0 | 0 | 0 | 5 | 10 | 1 |
| 變形菌門-γ綱 (Gammaproteobacteria) | 26 | 30 | 11 | 16 | 44 | 32 | 0 |
表3顯示了門水平上的相對微生物群落豐度。與對照相比,觀察到添加了電子供體的微觀體系中微生物群落組成發生了顯著變化。在所有添加了電子供體的微觀體系中,土壤中普遍存在的變形菌門和厚壁菌門是最豐富的門,它們在之前的鈾生物修復研究中也經常被觀察到。厚壁菌門在對照中的豐度遠低于在添加了電子供體的微觀體系中。在對照中檢測到放線菌門豐度較高,但在培養60天后,其豐度下降至不足0.5%。
總體而言,檢測到的科中有41個的相對豐度大于或等于1.0%。氣單胞菌科、假單胞菌科和梭菌科被檢測為葡萄糖、乙醇和乳酸添加的微觀體系中最豐富的科,而諾卡氏菌科和鞘脂單胞菌科是原始沉積物樣品中兩個相當豐富的科。在第90天的乳酸添加微觀體系中,根瘤菌科占10.3%,高于先前關于厭氧條件下地下水中U(VI)生物還原的研究。
