熱線:021-66110810,66110819
手機:13564362870
熱線:021-66110810,66110819
手機:13564362870
血細胞反應
在42%的分析蛤中記錄到強烈的血細胞浸潤。血細胞聚集主要出現在鰓絲內部和之間(圖6A)以及外套膜組織中(圖6B),盡管在內臟團中也觀察到血細胞浸潤。鰓中密集的血細胞聚集常常導致鰓結構的明顯破壞。血細胞反應偶爾與明顯的包涵體一起觀察到(圖7B),但更常見的是沒有發現病原體。
活蛤床和混合蛤床蛤類寄生蟲負荷的比較
不均勻的樣本量和低寄生蟲密度阻礙了對蛤床和混合床蛤類寄生蟲負荷進行統計學上穩健的比較。在兩種最流行的寄生蟲類型中,原生生物 I 在蛤床(39%)的流行率幾乎是混合床(23%)的兩倍。細菌感染在混合床(35%)比在蛤床(25%)更流行,組織血細胞浸潤也是如此(混合床:59%;蛤床:35%)。
大型生物
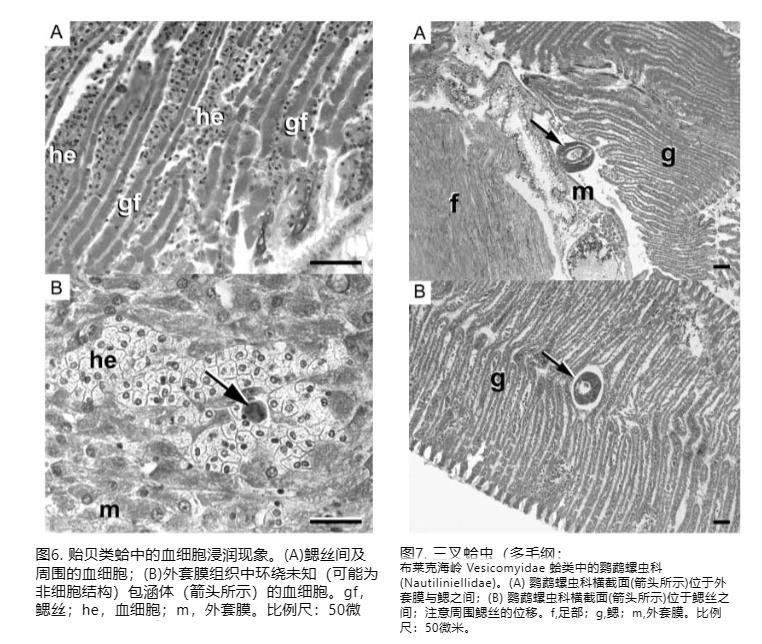
超過60%的受檢蛤含有一個或多個鸚鵡螺蟲多毛類。在大多數情況下,多毛類與蛤組織沒有密切關聯,但在三個實例中,幼年鸚鵡螺蟲與鰓組織緊密關聯,導致鰓絲移位(圖7)。
黑崖海丘囊蛤中的鸚鵡螺蟲多毛類(Nautiliniellidae)。(A) 鸚鵡螺蟲橫截面(箭頭)位于外套膜和鰓之間;(B) 鸚鵡螺蟲橫截面(箭頭)位于鰓絲之間;注意周圍鰓絲的移位。f,足;g,鰓;m,外套膜。比例尺:50微米。
討論
解釋滲口雙殼類大量死亡的任何嘗試都因我們無法知道死亡何時發生以及當時的環境條件如何而變得復雜。缺乏大量的沉積物覆蓋以及貝殼完整且未風化的情況表明,黑崖海丘現場的蛤類死亡相對較近(數月到數年,而不是數十年)。
海底硫化物通量的短暫變化已被認為是深海滲口環境中雙殼類大量死亡的原因(例如Jollivet, 1990; Van Dover等人,2003)。如果黑崖海丘蛤殼下方的沉積物氧化還原剖面與附近背景沉積物的氧化還原剖面相匹配,我們會得出結論,蛤中的化能自養共生體可能無法獲得足夠的硫化物生存。但是蛤殼床下方沉積物的還原條件表明,硫酸鹽還原和硫化物生成仍然強勁,饑餓在這種情況下不太可能是蛤類死亡的原因。然而,沒有硫化物濃度和通量的定量測量,我們不能確定棲息地質量未受硫化物毒性水平的損害。囊蛤的硫化物需求可能在同一物種內受到嚴格限制,并且在物種之間可變(Goffredi & Barry, 2002)。例如,蒙特雷灣冷泉共存的兩種囊蛤物種(Calyptogena pacifica 和 Calyptogena kilmeri)具有不同的硫化物吸收和同化模式(Goffredi & Barry, 2002)。Calyptogena pacifica 在硫化物吸收、運輸和氧化方面比 C. kilmeri 更高效,因此適合棲息在硫化物可用性較低的環境中(Goffredi & Barry, 2002)。C. kilmeri 及其共生體較低的硫化物代謝效率需要更高的環境硫化物水平(Goffredi & Barry, 2002)。即使硫化物可用性的微小變化也可能對蛤的健康和存活產生巨大影響(Goffredi & Barry, 2002),并可能解釋黑崖海丘現場的蛤類死亡。盡管對黑崖海丘囊蛤的具體硫化物需求存在不確定性,但蛤床、混合蛤殼床和蛤殼床下方相似的氧化還原剖面表明,除了流體通量變化之外的其他因素,包括寄生和疾病,可能促成囊蛤的死亡。
疾病對黑崖海丘雙殼類的潛在影響已在貽貝 Bathymodiolus heckerae 的組織病理學分析中得到證明(Ward等人,2004)。在這些貽貝的消化道中發現了一種致病性病毒樣包涵體,并與組織質量退化和繁殖能力下降相關(Ward等人,2004)。雖然疾病似乎在黑崖海丘貽貝種群結構中起作用(Ward等人,2004),但寄生對黑崖海丘蛤類的影響并不明顯。盡管在黑崖海丘貽貝(5種)和蛤類(6種)中觀察到的寄生蟲類型數量相似,但貽貝的感染密度和感染任何寄生蟲的個體數量都更高。消化組織通常是雙殼類的感染門戶(Lauckner, 1983; Pipe & Coles, 1995),其在囊蛤中的退化狀態(Boss & Turner, 1980; Goffredi & Barry, 2002)可能限制了病原體的侵入點。沒有一種蛤寄生蟲對整體組織質量產生重大影響。最常見的寄生蟲,原生生物 I,似乎是無害的。病毒樣包涵體 I(有時與組織壞死相關)和細菌感染 I(與鰓絲的局部破壞和退化相關),似乎是潛在的疾病因子,如果以更高的密度存在,可能促成這些種群的死亡。病毒感染與淺水軟體動物的嚴重死亡有關(Lauckner, 1983)。細菌感染 I 可能是繼發性定植者,利用因受傷或其他因素而受損的組織。可能的腸道細菌與消化上皮組織退化有關,但極低的感染流行率和密度,加上消化道在囊蛤營養中的相對不重要(Goffredi & Barry, 2002),表明這種寄生蟲不太可能在黑崖海丘滲口的蛤類群落中起結構作用。
迄今為止,已經使用光學顯微鏡描述了四種滲口軟體動物(兩種貽貝[Powell等人,1999; Ward等人,2004]、一種帽貝[Terlizzi等人,2004]和一種蛤[本研究])的微寄生蟲(表1)。在所有四種物種中都觀察到鰓和/或消化組織中的類立克次體包涵體和與鰓絲相關的原生生物(包括纖毛蟲)。病毒樣包涵體目前僅見于黑崖海丘的蛤類和貽貝。
| 宿主物種 地點 | 寄生蟲類型 | 感染部位 | 大小 | 參考文獻 |
|---|---|---|---|---|
|
Bathymodiolus sp. 墨西哥灣碳氫化合物滲口 550-560米 |
類立克次體菌落 | 鰓上皮 | 10-15微米 | Powell等人,1999 |
| 消化組織 | 25微米 | |||
| 細胞外鰓纖毛蟲 | 鰓絲之間 | 20微米 | ||
| 鰓玫瑰花結 | 鰓上皮 | 2-5微米細胞,每個液泡12至28個細胞 | ||
| 類Bucephalus包涵體 | 性腺(重度感染時全身) | 無數據 | ||
|
Bathymodiolus heckerae 黑崖海丘甲烷滲口 2155米 |
類立克次體菌落 細胞外鰓纖毛蟲 |
鰓和外套膜上皮 鰓絲之間 |
20微米 30微米 |
Ward等人,2004 |
| 類衣原體包涵體 | 消化組織 | 50微米 | ||
| 病毒樣包涵體 | 消化組織 | 5微米 | ||
|
Paralepetopsis floridensis 佛羅里達陡崖鹵水滲口 3288米 |
類立克次體菌落 | 消化組織 | 25-30微米 | Terlizzi等人,2004 |
| 細菌包涵體 | 消化組織 | 15微米 | ||
|
囊蛤 黑崖海丘甲烷滲口 2155米 |
類立克次體菌落 | 鰓上皮 | 20微米 8微米 | 本研究 |
| 可能的細菌包涵體 | 消化組織 | 2-3微米 | ||
| 細菌感染 | 鰓絲 | 200微米 | ||
| 病毒樣包涵體 | 鰓上皮 | 5微米 | ||
| 外套膜上皮 | 10微米 | |||
| 原生生物 | 鰓上皮 | 15微米 |
雖然幼年多毛類引起的鰓結構宏觀移位可能干擾鰓功能,但未發現與鸚鵡螺蟲多毛類 Vesicomyicola trifurcatus 相關的嚴重壞死。這些組織學觀察結果與鸚鵡螺蟲多毛類與其囊蛤宿主之間的共棲關系一致(Van Dover等人,2003)。
血細胞的吞噬作用是軟體的主要內部免疫防御(Pipe & Coles, 1995),血細胞浸潤可能是寄生蟲感染的跡象。大多數分析動物中存在血細胞反應,表明存在光學顯微鏡未檢測到的病原體。許多感染只能通過宿主反應的存在和強度來推斷(Lauckner, 1983),盡管血細胞浸潤并不總是疾病的指標;血細胞聚集也發生在對受傷或其他非感染性脅迫的反應中(Pipe & Coles, 1995)。
盡管在受檢蛤類中未檢測到強致病性寄生蟲,但存活率往往很少揭示疾病信息,因此現在將寄生排除為黑崖海丘蛤類種群死亡的潛在貢獻者為時過早,特別是因為從混合床收集的蛤類中細菌感染 I 和血細胞反應的流行率比從蛤床收集的蛤類高10%到20%。所有寄生蟲的感染密度都很輕,因此無法評估這些寄生蟲重度侵染的潛在影響;在低數量下看似無害的寄生蟲在較高密度下可能變得致病(Smolowitz, 1997)。例如,淺水雙殼類中的類 QPX 原生生物寄生蟲在健康個體中以低數量記錄,但在高密度存在時與嚴重死亡相關(Whyte等人,1994; Smolowitz, 1997)。
值得對黑崖海丘囊蛤的寄生進行進一步研究,應包括寄生蟲的電子顯微鏡研究,特別是病毒樣外套膜和鰓上皮包涵體,以確認感染因子的性質。使用特定染色靶向推定寄生蟲以及使用分子方法表征寄生蟲感染也是必要的。
