熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
輔酶 A 的三個(gè)磷酰基距離 Lys384 3.1 ?,并束縛著一個(gè)水分子網(wǎng)絡(luò)。幾個(gè)疏水殘基(Phe236、Met240 和 Ile316)定義了底物結(jié)合口袋的入口。輔酶 A 泛酸部分的 NH 基團(tuán)與 Ser130 主鏈的羰基在 2.7 ? 處形成氫鍵。Arg247 側(cè)鏈位于輔酶 A 泛酸部分內(nèi)的羰基之間。輔酶 A S 原子位于平面黃素基團(tuán)上方,距離其 3.1 ?,距離 Tyr243 OH 基團(tuán) 3.4 ?。Tyr243 的存在增加了空腔的極性,相比之下,在其他 Acds 中,相應(yīng)位置通常發(fā)現(xiàn)苯丙氨酸或亮氨酸。
脫輔基形式和類似物復(fù)合物晶體的晶胞內(nèi)容物不同,脫輔基形式中有一個(gè)二聚體,復(fù)合物結(jié)構(gòu)中有三個(gè)二聚體。兩種晶體都屬于相同的空間群 (P21212),并具有相同的 Matthews 系數(shù)。它們的晶胞尺寸通過 (0,1,0;3,0,0;0,0,1) 變換矩陣相關(guān)聯(lián)。在晶格中,這些變化通常是由于冷凍保護(hù)、冷卻和/或浸泡化合物的差異而發(fā)生的,這些化合物結(jié)合到晶體中并可能扭曲晶格,導(dǎo)致晶胞參數(shù)和空間群發(fā)生變化。在這種情況下,添加新鮮的琥珀酰輔酶 A 和從冷凍條件中省略甘油可能導(dǎo)致浸泡晶體的晶胞重排。Acd_DPN7 單體 A(殘基 2-392)在脫輔基形式和與輔酶 A 復(fù)合物的單體疊加顯示,僅外部環(huán)中的殘基 Gly143、Asp173 和 Gly190 的 r.m.s. 偏差(>1 ?)。Acd_DPN7 單體 A 和與輔酶 A 復(fù)合物的每個(gè)單體,在殘基 2-392 的 C 原子上計(jì)算的 r.m.s. 偏差 <0.5 ?。
3.4. Acd_DPN7 3SP-CoA 脫亞磺酰酶中 3SP-CoA 的建模
來自 D. multivorans 的戊二酰輔酶 A 脫氫酶與戊二酰輔酶 A 復(fù)合物的結(jié)構(gòu),以及來自豬肝線粒體的中鏈酰基輔酶 A 脫氫酶與辛酰輔酶 A 復(fù)合物的結(jié)構(gòu),與含有輔酶 A 基團(tuán)的 Acd_DPN7 3SP-CoA 脫亞磺酰酶結(jié)構(gòu)的疊加證實(shí),戊二酰輔酶 A 和辛酰輔酶 A 與各自脫氫酶的結(jié)合模式與輔酶 A 在 Acd_DPN7 空腔內(nèi)的結(jié)合模式相同。
為了研究活性位點(diǎn)中哪些殘基可能與底物的末端亞磺酰基相互作用,基于結(jié)合的輔酶 A 部分的結(jié)構(gòu)對(duì) 3SP-CoA 進(jìn)行了建模(圖 5a)。然后對(duì)模型進(jìn)行一輪分子動(dòng)力學(xué) (phenix.dynamics),使用默認(rèn)設(shè)置以消除模型偏差并獲得正確的幾何形狀。
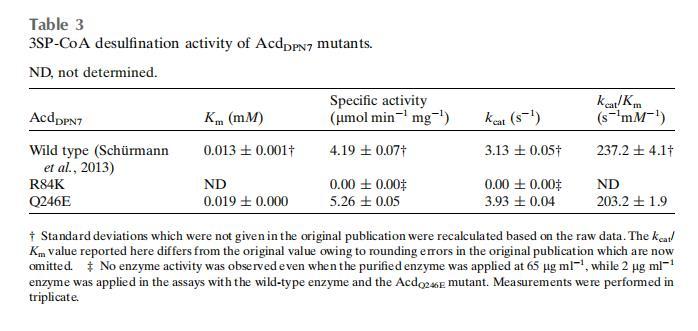
這清楚地顯示了丙酰基(C 原子位置 1-3;圖 3)的定位,亞磺酰基的 S 原子(位置 4)距離 Arg84、Asp88 和 Gln246 的側(cè)鏈在 3.5 ? 以內(nèi),位于 FAD 異咯嗪環(huán)平面的上方(圖 5a 和 5b)。一個(gè)水分子網(wǎng)絡(luò)可以容納在亞磺酰基的氫鍵距離內(nèi)(圖 5a)。
在這個(gè)模型中,3SP-CoA 硫酯鍵的羰基 O 原子既與 FAD 核糖醇側(cè)鏈的 2'-OH 形成氫鍵,也與 Gly366 的主鏈 NH 形成氫鍵。通常,在酰基輔酶 A 脫氫酶中,底物硫酯鍵的羰基 O 原子既與 FAD 核糖醇側(cè)鏈的 2'-OH 形成氫鍵,也與 Glu376 的主鏈 NH 形成氫鍵。這些相互作用對(duì)于底物羰基的取向和極化很重要。在脫氫酶的情況下,底物羰基的極化對(duì)于降低底物 α-碳的 pKa 值是必要的,這是脫氫反應(yīng)所必需的。這種極化效應(yīng)在 3SP-CoA 的脫亞磺酰反應(yīng)中是否也很重要,仍有待理解。
因此,Arg84、Asp88 和 Gln246 的側(cè)鏈以及 FAD 的異咯嗪環(huán)定義了脫亞磺酰反應(yīng)的酶促環(huán)境。3SP-CoA S4 與 FAD 異咯嗪環(huán)的距離將 >3.8 ?。
3.5. 突變酶 Acd_DPN7_R84K 和 Acd_DPN7_Q246E
Acd_DPN7 3SP-CoA 脫亞磺酰酶活性位點(diǎn)內(nèi) 3SP-CoA 的建模將亞磺酰基定位在靠近 Arg84 的位置,Arg84 NH2 與 3SP-CoA OS5 的距離為 3.2 ?(圖 5b)。殘基 Arg84 已被提議參與結(jié)合 3SP-CoA 的亞磺酰基,因?yàn)樵谖於]o酶 A 脫氫酶中觀察到了類似的相互作用。在這些酶中,一個(gè)精氨酸殘基參與結(jié)合末端羧基。為了研究該殘基對(duì)脫亞磺酰反應(yīng)是否必不可少,將 Arg84 突變?yōu)橘嚢彼幔a(chǎn)生 Acd_DPN7_R84K 變體。野生型 Acd_DPN7 和 Acd_DPN7_R84K 的尺寸排阻色譜表明,Acd_DPN7_R84K 被純化為完整的四聚體,因此是正確折疊的蛋白質(zhì)。Acd_DPN7_R84K 的吸收光譜與 Schürmann 等人發(fā)表的 Acd_DPN7 光譜相同。這表明所有 FAD 結(jié)合位點(diǎn)都已飽和,并且四聚體正確折疊。將 Arg84 突變?yōu)?Lys 導(dǎo)致活性完全喪失(表 3),表明 Arg84 具有重要作用。
亞磺酸比其對(duì)應(yīng)的羧酸酸性更強(qiáng),因此在生理 pH 下預(yù)計(jì)會(huì)去質(zhì)子化。Arg84 的胍基 pKa 為 12.5,預(yù)計(jì)在生理 pH 下帶正電荷,從而穩(wěn)定帶負(fù)電荷的亞磺酰基。考慮到氨基和胍基的 pKa 相似(Lys 和 Arg 分別為 11 和 12.5),預(yù)計(jì)兩個(gè)側(cè)鏈在生理 pH 下都會(huì)質(zhì)子化,并且都應(yīng)該能夠穩(wěn)定帶負(fù)電荷的亞磺酰基。Arg84 將帶有一個(gè)在胍基上離域的正電荷,而賴氨酸將在 NZ 原子上帶有一個(gè)非常局域化的正電荷。因此,電荷的穩(wěn)定可能只是 Arg84 側(cè)鏈作用的一個(gè)方面。此外,胍陽離子是強(qiáng)氫鍵供體。
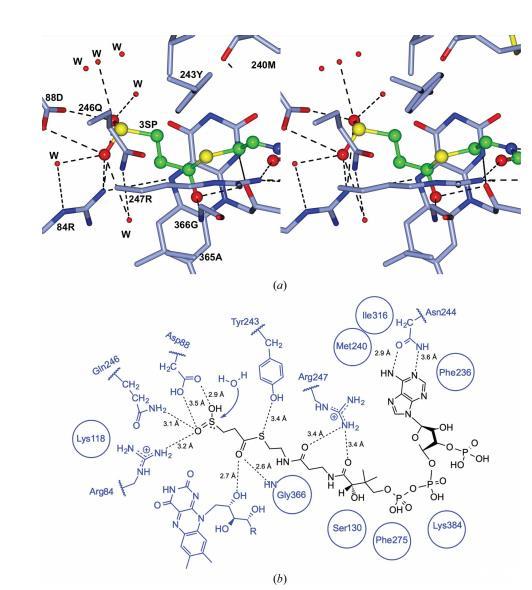
圖5 顯示了通過分子動(dòng)力學(xué)最小化處理后,A. mimigardefordensis來源的AcdDPN7(3SP-CoA脫硫酶)活性位點(diǎn)中3SP-CoA基團(tuán)的定位視圖。3SP-CoA的定位基于3SP-CoA脫硫酶腔內(nèi)CoA的晶體學(xué)精修位置。(a)腔深端立體視圖。3SP-CoA以球棍模型表示,原子顏色編碼如下:碳原子綠色、氧原子紅色、氮原子藍(lán)色、硫原子黃色。蛋白質(zhì)以青色粗鍵表示,其中氧原子為紅色、氮原子為藍(lán)色。水分子(W)以紅色球體表示。(b)整個(gè)結(jié)合位點(diǎn)的二維表示。
在模型中,Gln246 NE2 與 3SP-CoA OS5 的距離為 3.1 ?(圖 5b),并且先前假設(shè) Gln246 的存在是 Acd_DPN7 無法催化酰基輔酶 A 脫氫酶反應(yīng)的原因。因此,產(chǎn)生了 Acd_DPN7_Q246E 變體。該變體仍然能夠作為脫亞磺酰酶(表 3),但不能利用任何測(cè)試的酰基輔酶 A 脫氫酶底物(丙酰輔酶 A、丁酰輔酶 A、異丁酰輔酶 A、戊酰輔酶 A、異戊酰輔酶 A、琥珀酰輔酶 A、戊二酰輔酶 A 和 3SP-CoA)(結(jié)果未顯示)。因此,Acd_DPN7_Q246E 突變體在谷氨酰胺突變?yōu)楣劝彼岷蟛⑽传@得酰基輔酶 A 脫氫酶活性。然而,在 Acd_DPN7_Q246E 突變體中,取代對(duì)脫亞磺酰速率有輕微影響(表 3)。Km 從野生型到突變體增加了約 46%,而 kcat 值增加了約 25%。底物結(jié)合似乎部分受損,可能是由于新添加的帶負(fù)電荷的羧基(谷氨酰胺突變?yōu)楣劝彼幔@可能會(huì)排斥帶負(fù)電荷的 3-亞磺酰丙酰基的進(jìn)入。反應(yīng)速率可能因空腔帶負(fù)電荷更多而增強(qiáng),這將有助于在脫亞磺酰后去除 HSO3 基團(tuán)。
 相關(guān)新聞
相關(guān)問題
相關(guān)新聞
相關(guān)問題