熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
Diverse electron sources support denitrification under hypoxia in the obligate methanotroph Methylomicrobium album strain BG8
不同的電子源支持專性嗜甲烷菌 Methylomicrobium album 菌株 BG8 在缺氧下的反硝化作用
來源:Front. Microbiol. 6:1072.
論文摘要
本研究探討了專性嗜甲烷菌 Methylomicrobium album菌株 BG8 在缺氧條件下利用多種電子源支持反硝化作用的能力。研究發(fā)現(xiàn),BG8 菌株能夠利用甲烷、甲醇、甲醛、甲酸、乙烷、乙醇和氨作為電子供體,在亞硝酸鹽(NO??)存在下于缺氧環(huán)境中驅(qū)動反硝化過程,產(chǎn)生氧化亞氮(N?O)。轉(zhuǎn)錄組分析顯示,反硝化相關(guān)基因(如 nirS和 norB1)的表達(dá)在 NO?? 添加后顯著上調(diào),同時一個推測的銅單加氧酶基因 pxmA的表達(dá)也受低氧和 NO?? 誘導(dǎo)。結(jié)果表明,嗜甲烷菌通過表達(dá)反硝化基因庫,能夠在氧限制下將底物氧化與氮氧化物還原偶聯(lián),擴(kuò)展了其對環(huán)境變化的代謝靈活性認(rèn)知。
研究目的
本研究旨在:
驗證 M. albumBG8 是否能利用多種電子源(包括 C1 化合物、C2 化合物和無機(jī)氮源)在缺氧條件下支持反硝化活性。
確定反硝化過程的環(huán)境調(diào)控因素,特別是氧張力和 NO?? 可用性的作用。
評估 BG8 菌株反硝化基因(如 nirS、norB)和潛在新基因(如 pxmA)的表達(dá)響應(yīng)。
闡明嗜甲烷菌在氮循環(huán)中的生態(tài)角色,尤其是其對 N?O 排放的潛在貢獻(xiàn)。
研究思路
研究采用多方法結(jié)合的實驗策略:
菌株培養(yǎng)與處理:在硝酸鹽礦物鹽培養(yǎng)基(NMS)中培養(yǎng) BG8 菌株,設(shè)置對照(僅 NMS)和實驗組(添加 1 mM NaNO?),在控制氧張力(初始約 21% O?)下監(jiān)測 120 小時。
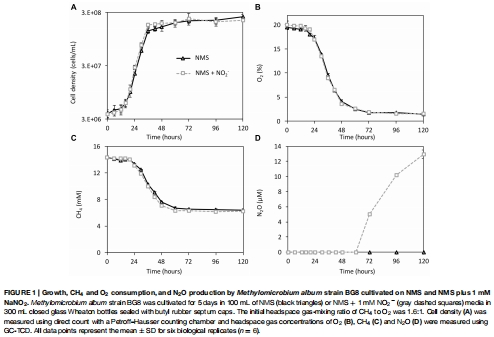
氣體動態(tài)監(jiān)測:使用氣相色譜(GC-TCD)定期測量培養(yǎng)瓶頂空中的 O?、CH? 和 N?O 濃度,評估生長、底物消耗和反硝化產(chǎn)物生成(圖1)。
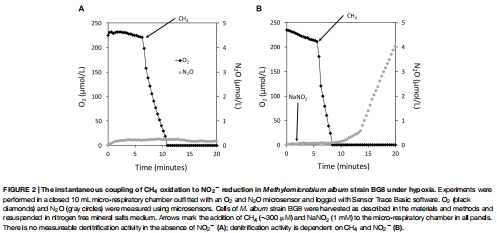
微傳感器瞬時測量:在微呼吸室(10 mL)中,使用丹麥Unisense O? 微電極和N?O 微傳感器同步監(jiān)測休眠細(xì)胞在添加不同電子源(如 CH?、CH?OH、NH?Cl 等)后的溶解 O? 消耗和 N?O 生成速率(圖2、3、4)。
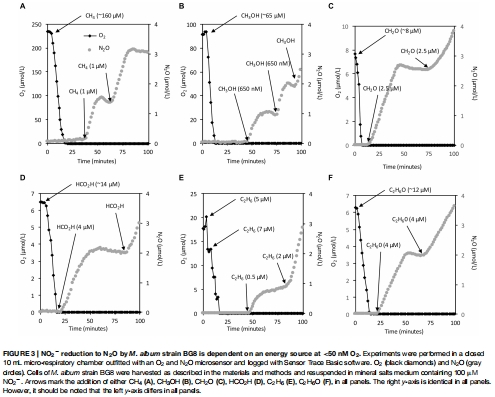
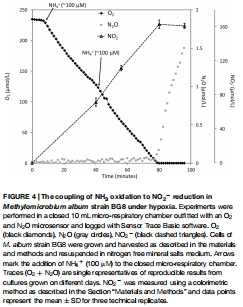
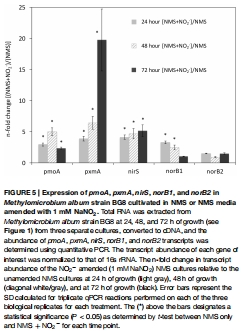
基因表達(dá)分析:通過 RNA 提取和定量 PCR(qPCR)測量反硝化基因(nirS、norB1、norB2)和代謝基因(pmoA、pxmA)的轉(zhuǎn)錄水平,比較 NO?? 添加組與對照組的差異(圖5)。
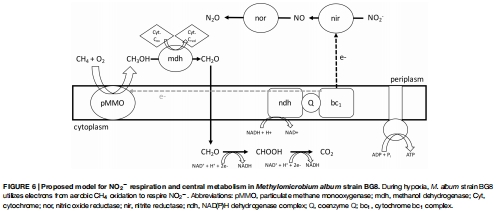
數(shù)據(jù)整合:將氣體通量數(shù)據(jù)與基因表達(dá)關(guān)聯(lián),構(gòu)建反硝化代謝模型(圖6)。
測量的數(shù)據(jù)、研究意義及來源
研究測量了多類數(shù)據(jù),以下按類別說明其意義及圖表來源(注明圖表):
生長與氣體代謝數(shù)據(jù):
數(shù)據(jù)內(nèi)容:BG8 在添加 NO?? 后生長不受抑制,但 O? 消耗加速,且在 O? 降至約 1.8% 時開始產(chǎn)生 N?O(72 小時);N?O 產(chǎn)率達(dá) 5.1% 從添加的 NO??(100 μmol)。無 NO?? 組無 N?O 檢測。
研究意義:直接證明反硝化嚴(yán)格依賴缺氧和 NO??,且不影響菌株正常生長,凸顯 BG8 的代謝穩(wěn)健性。數(shù)據(jù)來自圖1A-D(生長曲線和氣體變化)。
微傳感器瞬時O?與N?O通量數(shù)據(jù):
數(shù)據(jù)內(nèi)容:在微呼吸室中,添加 CH? 后 O? 在 3 分鐘內(nèi)耗盡,隨后 NO?? 還原產(chǎn)生 N?O(速率 7.9 × 10?1? mol cell?1 h?1)。類似趨勢見于其他底物(如甲醇、乙烷、氨),N?O 生成與底物添加量成正比。
研究意義:提供電子源與反硝化活性的直接耦合證據(jù),表明多種底物可通過共同酶學(xué)途徑驅(qū)動氮氧化物還原。數(shù)據(jù)來自圖2A-F(各底物下的 O?/N?O 曲線)和圖3(底物劑量響應(yīng))。
氨氧化耦合反硝化數(shù)據(jù):
數(shù)據(jù)內(nèi)容:添加 NH?? 后,O? 快速消耗,NO?? 積累至 163 μM,缺氧后 N?O 以 1.2 × 10?1? mol cell?1 h?1 速率生成。
研究意義:揭示無機(jī)氮源(氨)也可支持反硝化,擴(kuò)展了嗜甲烷菌的能量獲取策略。數(shù)據(jù)來自圖4(NH?? 添加實驗)。
基因表達(dá)數(shù)據(jù)(qPCR):
數(shù)據(jù)內(nèi)容:NO?? 添加組中 nirS和 norB1轉(zhuǎn)錄水平在 24-48 小時顯著升高(如 nirS增加 4.5 倍),pxmA在 72 小時上調(diào) 19.8 倍;pmoA(甲烷氧化基因)表達(dá)穩(wěn)定。
研究意義:從分子層面證實反硝化基因?qū)?NO?? 的特異性響應(yīng),提示 pxmA可能參與缺氧適應(yīng)。數(shù)據(jù)來自圖5(基因表達(dá)柱狀圖)。
研究結(jié)論
M. albumBG8 在缺氧和 NO?? 存在下能進(jìn)行反硝化,產(chǎn)生 N?O,且過程依賴多種電子源(C1/C2 化合物及氨),體現(xiàn)其顯著的代謝可塑性。
反硝化活性由基因表達(dá)調(diào)控驅(qū)動:nirS和 norB1對 NO?? 響應(yīng)靈敏,而 pxmA的上調(diào)暗示其可能參與新型缺氧應(yīng)激途徑。
嗜甲烷菌的反硝化能力可能影響自然環(huán)境中氮循環(huán),尤其在氧限制生境(如濕地、沉積物)中貢獻(xiàn)于 N?O 排放。
研究為工程化利用嗜甲烷菌于溫室氣體減排(如 CH? 和 N?O 協(xié)同控制)提供了理論基礎(chǔ)。
詳細(xì)解讀使用丹麥Unisense電極測量出來的數(shù)據(jù)的研究意義
在本文中,使用丹麥Unisense O? 微電極和N?O 微傳感器測量的數(shù)據(jù)具有關(guān)鍵研究意義,其詳細(xì)解讀如下:
高時空分辨率實時監(jiān)測代謝動態(tài):Unisense 微傳感器是一種電化學(xué)傳感器,具備高靈敏度(檢測限達(dá)納摩爾級)和快響應(yīng)時間(秒級)。本研究將其集成于微呼吸室,以秒級間隔同步測量溶解 O? 和 N?O 的濃度變化(如圖2A顯示 O? 在 3 分鐘內(nèi)耗盡)。這種實時原位監(jiān)測能力避免了傳統(tǒng)終點取樣的時間模糊性,直接捕捉了反硝化啟動的精確時刻(O? 耗盡后立即發(fā)生),為“缺氧觸發(fā)反硝化”的假設(shè)提供了動態(tài)證據(jù)。
定量驗證電子源與反硝化的直接耦合:通過添加不同底物(如 CH?、CH?OH、NH??)后記錄 O? 消耗和 N?O 生成曲線(圖2-4),Unisense 數(shù)據(jù)將抽象的“電子供體”轉(zhuǎn)化為可量化的通量速率。例如,數(shù)據(jù)顯示所有測試底物均能支持相似的 N?O 產(chǎn)率,證明 BG8 的反硝化酶系具有廣譜底物適應(yīng)性。這種精確量化是推導(dǎo)代謝模型(圖6)的基礎(chǔ)。
揭示反硝化的瞬時調(diào)控機(jī)制:微傳感器數(shù)據(jù)顯示,N?O 生產(chǎn)僅在 O? 降至檢測限(<50 nM)后開始(圖2B),強(qiáng)有力地證實了反硝化對缺氧的絕對依賴性。同時,O? 消耗速率與底物類型相關(guān)(如 CH? 最快),將電子流效率與反硝化活性關(guān)聯(lián),支持了“反硝化作為缺氧備用呼吸鏈”的生理模型。
方法學(xué)優(yōu)勢與可靠性:Unisense 傳感器的原位校準(zhǔn)(用無氧和空氣飽和溶液)和微創(chuàng)測量確保了數(shù)據(jù)準(zhǔn)確性。在本文中,其與 GC 數(shù)據(jù)互補(bǔ)(如圖1),通過高時間分辨率驗證了批量培養(yǎng)的整體趨勢,凸顯了微傳感器在解析瞬時微生物過程中的不可替代性。
總之,丹麥Unisense電極的數(shù)據(jù)不僅是本研究中驗證反硝化活性的技術(shù)基石,更通過提供高精度實時通量信息,揭示了嗜甲烷菌在缺氧下的代謝切換機(jī)制,為理解微生物在碳氮耦合循環(huán)中的功能提供了關(guān)鍵見解。