熱線:021-66110810,66110819
手機(jī):13564362870
熱線:021-66110810,66110819
手機(jī):13564362870
Post-metamorphic carryover effects of larval digestive plasticity
幼蟲消化可塑性的后變質(zhì)延續(xù)效應(yīng)
來(lái)源:Animal Physiological Ecology doi: 10.1111/1365-2435.12501
論文摘要
本研究以紅眼樹蛙(Agalychniscallidryas)為模型,探究了具有復(fù)雜生命周期的動(dòng)物,其幼蟲期環(huán)境條件(種內(nèi)競(jìng)爭(zhēng))是否會(huì)對(duì)變態(tài)后的生理和生長(zhǎng)產(chǎn)生持續(xù)效應(yīng)。研究發(fā)現(xiàn),在低、中、高三種幼蟲密度下,高密度環(huán)境誘導(dǎo)了幼蟲的消化可塑性,表現(xiàn)為腸道更長(zhǎng),但肝臟和脂肪體更小。這些效應(yīng)在變態(tài)后依然持續(xù):來(lái)自高幼蟲密度的幼蛙體型更小,但具有相對(duì)更長(zhǎng)的腸道和極小的肝臟與脂肪體。此外,幼蟲密度差異還導(dǎo)致了變態(tài)后攝食行為的顯著不同:來(lái)自高密度的小型幼蛙在變態(tài)完成前即開始攝食,而來(lái)自低密度的大型幼蛙則延遲約兩周才開始攝食。一旦開始攝食,不同體型幼蛙的攝入量和食物平均留存時(shí)間無(wú)顯著差異。研究表明,幼蟲期的消化可塑性及其對(duì)能量?jī)?chǔ)備(肝臟、脂肪體)的持續(xù)影響,是驅(qū)動(dòng)幼蛙出現(xiàn)體型依賴性生長(zhǎng)模式(先前報(bào)道的小型幼蛙生長(zhǎng)更快)的關(guān)鍵機(jī)制。
研究目的
本研究旨在驗(yàn)證以下核心假設(shè)與問(wèn)題:
幼蟲期的種內(nèi)競(jìng)爭(zhēng)(密度)是否會(huì)對(duì)紅眼樹蛙幼蟲的內(nèi)部器官(腸道、肝臟、脂肪體)大小產(chǎn)生可塑性影響。
這種幼蟲期的器官可塑性是否會(huì)持續(xù)到變態(tài)后階段,盡管兩棲動(dòng)物變態(tài)伴隨著腸道從植食性到肉食性的劇烈重塑。
幼蟲密度差異是否會(huì)導(dǎo)致幼蛙在變態(tài)后的攝食行為(如開始攝食的時(shí)間、攝入率)和代謝率產(chǎn)生差異。
探究幼蟲期的消化可塑性能否解釋該物種中觀察到的“小型幼蛙生長(zhǎng)更快”的現(xiàn)象。
研究思路
研究采用對(duì)照實(shí)驗(yàn)與持續(xù)觀測(cè)相結(jié)合的方法:
幼蟲飼養(yǎng)與分組:在戶外中宇宙環(huán)境中,將紅眼樹蛙幼蟲分別置于低(5只/缸)、中(25只/缸)、高(50只/缸)三種密度下飼養(yǎng)。
幼蟲期樣本采集與測(cè)量:在幼蟲發(fā)育到特定大小時(shí),取樣并解剖。測(cè)量其外部形態(tài)(體長(zhǎng)、體重)和內(nèi)部器官(腸道長(zhǎng)度和質(zhì)量、肝臟質(zhì)量、脂肪體質(zhì)量)。
變態(tài)后樣本采集與測(cè)量:收集從各密度組變態(tài)而來(lái)的幼蛙,測(cè)量其體型(體長(zhǎng)、體重),并在尾巴吸收完成后一天進(jìn)行解剖,測(cè)量其腸道、肝臟和脂肪體的大小。
攝食行為觀測(cè):跟蹤觀察幼蛙變態(tài)后開始產(chǎn)生糞便的時(shí)間、每日糞便中的獵物數(shù)量(以評(píng)估攝入量),并通過(guò)一次性投喂標(biāo)記性食物(葉蟬)來(lái)測(cè)量食物的平均留存時(shí)間。
代謝率測(cè)量:使用丹麥Unisense封閉系統(tǒng)呼吸計(jì)測(cè)量幼蛙(在未攝食狀態(tài)下)的耗氧率,作為其代謝率的指標(biāo)。
數(shù)據(jù)分析:使用方差分析、協(xié)方差分析等統(tǒng)計(jì)方法,比較不同幼蟲密度組在幼蟲期和變態(tài)后各指標(biāo)的差異。
測(cè)量的數(shù)據(jù)、研究意義及來(lái)源
研究測(cè)量了多方面的數(shù)據(jù):
幼蟲器官形態(tài)數(shù)據(jù):
數(shù)據(jù)內(nèi)容:高密度幼蟲的腸道質(zhì)量和長(zhǎng)度顯著大于低密度幼蟲,但其肝臟和脂肪體質(zhì)量顯著更小。
研究意義:直接證明了幼蟲密度(種內(nèi)競(jìng)爭(zhēng))能誘導(dǎo)消化器官(腸道增長(zhǎng)以提升在低質(zhì)量環(huán)境中的消化效率)和能量?jī)?chǔ)存器官(肝臟、脂肪體減小,反映能量狀況較差)的可塑性變化。這些數(shù)據(jù)來(lái)自表1。
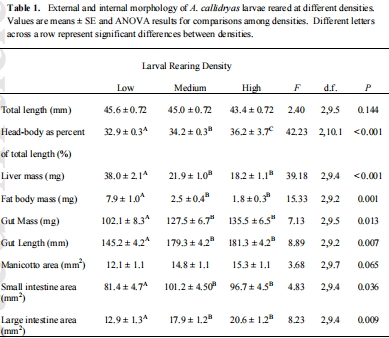
幼蛙器官形態(tài)數(shù)據(jù):
數(shù)據(jù)內(nèi)容:盡管來(lái)自高密度組的幼蛙體型顯著小于低密度組,但其腸道長(zhǎng)度和質(zhì)量并無(wú)差異,導(dǎo)致其相對(duì)腸道長(zhǎng)度更長(zhǎng);而肝臟和脂肪體大小隨幼蟲密度增加而顯著減小。
研究意義:證明了幼蟲期的器官可塑性(尤其是腸道延長(zhǎng)和能量?jī)?chǔ)備減少)能夠持續(xù)到變態(tài)后階段,盡管經(jīng)歷了劇烈的腸道重塑。這是“持續(xù)效應(yīng)”的直接形態(tài)學(xué)證據(jù)。數(shù)據(jù)來(lái)自表2。
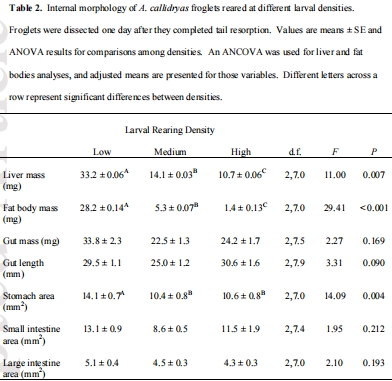
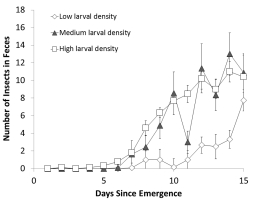
攝食行為數(shù)據(jù):
開始攝食時(shí)間:來(lái)自高幼蟲密度的幼蛙開始產(chǎn)生糞便的時(shí)間(平均5.7天)顯著早于低密度組(平均8.0天),且87%的高密度組個(gè)體在尾巴吸收完成前就已開始攝食。
早期攝入量:在變態(tài)后早期,來(lái)自高密度組的幼蛙攝食率更高。
研究意義:表明幼蟲期的能量狀況(低能量?jī)?chǔ)備)驅(qū)動(dòng)了變態(tài)后的攝食行為,能量?jī)?chǔ)備低的個(gè)體更早、更積極地開始攝食以補(bǔ)償能量不足。這些數(shù)據(jù)來(lái)自圖2及相關(guān)文本描述。
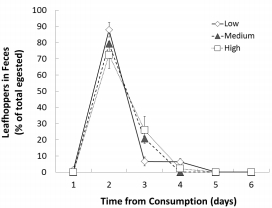
食物平均留存時(shí)間數(shù)據(jù):
數(shù)據(jù)內(nèi)容:一次性投喂葉蟬后,葉蟬頭殼主要在第二天出現(xiàn)在糞便中。計(jì)算出的平均留存時(shí)間約為2.22天,且在不同幼蟲密度組間無(wú)顯著差異。
研究意義:表明盡管攝入時(shí)間和攝入量有差異,但幼蛙的基本消化效率(食物通過(guò)腸道的時(shí)間)可能相似,排除了消化效率差異是生長(zhǎng)模式主因的可能性。數(shù)據(jù)來(lái)自圖3。
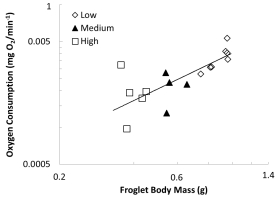
使用丹麥Unisense電極測(cè)量的代謝率數(shù)據(jù):
數(shù)據(jù)內(nèi)容:幼蛙的代謝率與體重呈顯著正相關(guān)(異速生長(zhǎng)斜率約為1.16),但在控制了體重因素后,不同幼蟲密度組間的體重特異性代謝率無(wú)顯著差異。
研究意義:表明幼蟲密度導(dǎo)致的持續(xù)效應(yīng)并未顯著影響幼蛙的基礎(chǔ)代謝率。生長(zhǎng)模式的差異不能歸因于能量消耗的差異,從而將原因指向了能量攝入端(攝食行為)和能量分配策略。數(shù)據(jù)來(lái)自圖4。
研究結(jié)論
顯著的持續(xù)效應(yīng):幼蟲期的種內(nèi)競(jìng)爭(zhēng)對(duì)紅眼樹蛙的形態(tài)(腸道、能量?jī)?chǔ)備器官)產(chǎn)生了可塑性影響,并且這些影響持續(xù)到了變態(tài)后階段。
能量?jī)?chǔ)備驅(qū)動(dòng)攝食行為:來(lái)自高幼蟲密度的幼蛙能量?jī)?chǔ)備(肝臟、脂肪體)極低,這促使它們更早、更積極地開始攝食,表現(xiàn)出補(bǔ)償生長(zhǎng)的傾向。
代謝率無(wú)差異:幼蟲密度不影響變態(tài)后幼蛙的體重特異性代謝率,說(shuō)明能量消耗不是導(dǎo)致生長(zhǎng)差異的主要原因。
機(jī)制解釋:幼蟲期誘導(dǎo)的更長(zhǎng)腸道(可能提升消化能力)coupledwith更早更積極的攝食行為(以補(bǔ)償能量赤字),共同解釋了為何小型幼蛙(來(lái)自高密度)在變態(tài)后早期生長(zhǎng)更快。這是一種應(yīng)對(duì)不利幼蟲環(huán)境的適應(yīng)性策略。
詳細(xì)解讀使用丹麥Unisense電極測(cè)量出來(lái)的數(shù)據(jù)的研究意義
使用丹麥Unisense呼吸測(cè)量系統(tǒng)(配備光纖氧傳感器)測(cè)量的耗氧量數(shù)據(jù)在本研究中具有關(guān)鍵的研究意義,其詳細(xì)解讀如下:
精確量化基礎(chǔ)代謝率:Unisense系統(tǒng)是一種高精度的封閉式呼吸計(jì),能夠?qū)崟r(shí)測(cè)量密閉腔體內(nèi)氧氣分壓的下降速率。本研究在幼蛙未攝食的狀態(tài)下測(cè)量其耗氧率,這有效地反映了動(dòng)物的標(biāo)準(zhǔn)代謝率,即維持基本生命活動(dòng)所需的最小能量消耗。這為比較不同處理組動(dòng)物的能量消耗提供了可靠、可比較的基線數(shù)據(jù)。
排除代謝差異作為生長(zhǎng)差異的驅(qū)動(dòng)因素:測(cè)量得到的數(shù)據(jù)顯示,代謝率與體重顯著相關(guān)(圖4),但最關(guān)鍵的是,在統(tǒng)計(jì)上控制了體重的影響后,不同幼蟲密度來(lái)源的幼蛙其體重特異性代謝率沒有顯著差異。這一發(fā)現(xiàn)具有重要的排除性意義:它強(qiáng)有力地表明,觀察到的生長(zhǎng)模式差異(小個(gè)體生長(zhǎng)更快)并非由于它們具有更高的代謝效率或更低的維持能耗所致。如果小個(gè)體代謝率顯著更高或更低,都會(huì)使機(jī)理解釋復(fù)雜化。此數(shù)據(jù)將生長(zhǎng)差異的原因清晰地指向了能量攝入端(攝食行為)和能量分配策略,而非能量消耗端。
支持“補(bǔ)償生長(zhǎng)”的能量分配假說(shuō):該數(shù)據(jù)支持了以下解釋:來(lái)自高密度的小型幼蛙,雖然基礎(chǔ)代謝需求與同等體重的個(gè)體相似,但它們通過(guò)更早開始攝食和可能更高的攝入率,將更多的凈能量用于生長(zhǎng),以補(bǔ)償幼蟲期的發(fā)育滯后。而來(lái)自低密度的大型幼蛙,由于能量?jī)?chǔ)備相對(duì)充足,不急于攝食,將能量?jī)?yōu)先用于維持或其他活動(dòng),生長(zhǎng)投入相對(duì)較少。Unisense數(shù)據(jù)證實(shí)了這種不同的能量分配策略是發(fā)生在攝入能量之后,而不是由不同的基礎(chǔ)代謝率所預(yù)先決定的。
方法學(xué)可靠性:該系統(tǒng)校準(zhǔn)嚴(yán)格(使用水飽和空氣和無(wú)氧水),測(cè)量環(huán)境控溫,確保了數(shù)據(jù)的準(zhǔn)確性和可比性,為上述核心結(jié)論提供了堅(jiān)實(shí)的技術(shù)支撐。
綜上所述,丹麥Unisense電極測(cè)量的代謝率數(shù)據(jù)在本研究中起到了“排除法”的關(guān)鍵作用。它通過(guò)精確證實(shí)“代謝率無(wú)組間差異”,有效地排除了能量消耗差異這一潛在解釋,從而將研究的焦點(diǎn)和最終結(jié)論引導(dǎo)至攝食行為可塑性和能量分配策略上,深化了對(duì)幼蟲期環(huán)境如何通過(guò)持續(xù)效應(yīng)影響變態(tài)后生活史策略的理解。